|
Research field and expertise
Marine ecology, Marine biology, Fisheries biology, Fisheries oceanography
Small pelagic fish, Population dynamics, Recruitment mechanism, Growth and survival, Early life history, Spawning biology
Main themes
Biological mechanisms of species alternations
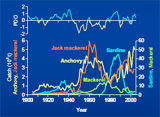
Marine ecosystems are notorisously complex. Despite such a nature, small pelagic fish have exhibited cyclic patterns of population dynamics worlwide. A typical example is the phenomenon of out-of-phase population oscillations between anchovy and sardine. These species alternations have been associated with climate changes; however, the biological processes have remained unresolved. We have been exploring potential mechanisms to explain the biological processes of multi-species regime shifts of small pelagic fish.
Key questions:
- Why do even subtle environmental changes sometimes trigger dramatic fish regime shifts?
- Why do anchovy flourish and sardine collapse or vice versa under the same ocean regime?
- Why are anchovy and sardine alternations synchronous or asynchronous among different ecosystems?
Key contents:
- "Optimal growth temperature" hypothesis (Takasuka & Aoki 2006 in FO, Takasuka et al. 2007 in CJFAS)
- Contrasting spawning temperature optima (Takasuka et al. 2008 in PO)
- Multi-species regime shifts (Takasuka et al. 2008 in MEPS)
- Spawning overlap (Takasuka et al. 2008 in MEPS)
- Characterizing spawning habitats (Oozeki et al. 2007 in CalCOFI)
- Expansion/contraction (Barange et al. 2009 in PO)
- Mechanism review (Drinkwater et al. 2009 in JMS)
Growth-based survival mechanisms during early life stages
Fish experience severe challenges to survival during the early life stages. Growth rates dominate survival potential suring those stages, playing a key role in survival dynamics. The "growth-survival" paradigm posits that faster growing individuals will have a higher probability of survival than slower growing conspecifics. This theory had been explained indirectly by two concepts: size and time (the "bigger is better" and "stage duration" hypotheses, respectively). Predation is the major and direct source of mortality. Nonetheless, no direct evidence had yet been obtained to support the relationship between growth rates and predation mortality. In our field studies, larval anchovy and their potential predators were captured simultaneously by the same tows. Through otolith microstructure analysis, we directly examined growth rates of the ingested larvae from the stomach contents of predatory fish and compared them with those of the surviving larvae from the original populations to demonstrate direct impacts of growth rates on predation mortality.
Key questions:
- Is a slower growing fish larva actually removed by predation at a given moment in the sea?
- If so, is the mortality size-dependent?
- Is there any difference among predatory species?
Key contents:
- "Growth-selective predation" hypothesis (Takasuka et al. 2003 in MEPS)
- Three synergistic growth-related mechanisms (Takasuka et al. 2004 in MEPS)
- "Growth-selective predation" hypothesis revisited (Takasuka et al. 2004 in MEPS)
- Predator-specific "growth-selective predation" (Takasuka et al. 2007 in MEPS)
- Growth effect on the otolith and somatic sizes (Takasuka et al. 2008 in FS)
- Growth in the Kii Channel (Yasue & Takasuka 2009 in JFB)
- Predation dynamics of mackerel (Robert et al. 2010)
- Dynamics of growth-based survival mechanisms (Takasuka et al. submitted)
Specific keywords
"Growth-selective predation" mechanism A mechanism linking growth rate to predation mortality (Takasuka et al. 2003 in MEPS) under the theoretical framework of the "growth-survival" paradigm. Slower growing larvae are more vulnerable to predation mortality than faster growing conspecifics, even if they are the same size (i.e. non-size-related), at a given moment (i.e. non-time-related) in the sea. This means that the level of growth rate per se has direct impacts on vulnerability to predation, independently of both size (negative size-selective mortality) and time (stage duration). We proposed the "growth-selective predation" hypothesis (mechanism), which is theoretically independent of and synergistic with the existing hypotheses based on size and time ("bigger is better" and "stage duration" hypotheses, respectively) under a general concept of the "growth-mortality" hypothesis.
The first demonstration was conducted by direct comparison of growth rates between the larvae actually ingested by the predators and surviving larvae from the original populations in Sagami Bay (Takasuka et al. 2003 in MEPS). The hypothesis was then tested based on the characteristics of the survivors versus original populations (Takasuka et al. 2004 in MEPS) and revisited for larval anchovy in offshore waters (Takasuka et al. 2004 in MEPS). A further study demonstrated predator-specific "growth-selective predation" on larval anchovy with multiple samples (Takasuka et al. 2007 in MEPS).
"Optimal growth temperature" hypothesis A potential biological mechanism for anchovy and sardine alternations (Takasuka et al. 2007 in CJFAS). A mystery of the ocean is the phenomenon of out-of-phase population oscillations between anchovy and sardine. Why do even subtle environmental changes sometimes trigger dramatic alternations? Why do anchovy flourish and sardine collapse or vice versa under the same ocean regime? We propose a simple "optimal growth temperature" hypothesis, in which anchovy and sardine alternations are caused by differential optimal temperatures for growth rates during the early life history stages. The "growth-survival" paradigm, direct temperature impacts, and differential optimal temperatures for growth rates constitute the bottom line of this concept. Subsequently, the hypothesis was extended to the multi-species regime shifts of small pelagic fish in the western North Pacific (Takasuka et al. 2008 in MEPS) and synchronous alternations during certain phases between the opposite sides of the North Pacific (Takasuka et al.2008 in PO), focusing on species-specific spawning temperature optima.
The website of current researches by Akinori Takasuka (¹³É{ēĻ¢ŠōT/ŹéÉ{ēĻ¢ŠōT).